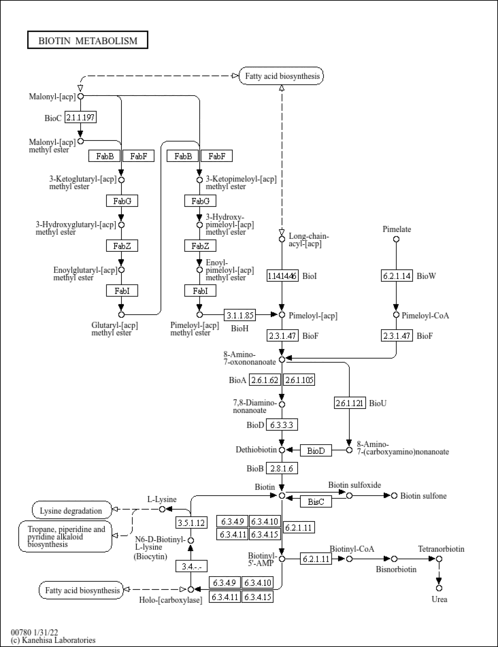
Biotin (vitamin H or vitamin B7) is the essential cofactor of biotin-dependent carboxylases, such as pyruvate carboxylase and acetyl-CoA carboxylase. Mammals cannot synthesize biotin, while in bacteria, fungi, and plants it is synthesized from pimelate thioester through different pathways. In E. coli and many organisms, pimelate thioester is derived from malonyl-ACP. The pathway starts with the methylation to malonyl-ACP methyl ester, followed by the fatty acid chain elongation cycle to form pimeloyl-ACP methyl ester, which is then demethylated to form pimeloyl-ACP [MD:M00572]. Pimeloyl-ACP is converted to biotin through the final four steps in the biotin bicyclic ring assembly, which are conserved among biotin-producing organisms [MD:M00123]. In B. subtilis, biotin is derived from pimeloyl-ACP formed by oxidative cleavage of long-chain acyl-ACPs [MD:M00573]. Some bacteria synthesize biotin from pimeloyl-CoA derived from pimelate [MD:M00577]. Biotin is covalently attached to biotin-dependent carboxylase by biotin protein ligase, also known as holocarboxylase synthase in mammals, to form an active holocarboxylase. After degradation of the biotinylated carboxylase into biocytin, it is further degraded by biotinidase to release free biotin, which is recycled in holocarboxylase synthesis. Biotin is catabolized by beta-oxidation of the valeric acid side chain or oxidation of sulfur in the heterocyclic ring.
Expression, purification, and characterization of BioI: a carbon-carbon bond cleaving cytochrome P450 involved in biotin biosynthesis in Bacillus subtilis.
Ploux O, Soularue P, Marquet A, Gloeckler R, Lemoine Y
Title
Investigation of the first step of biotin biosynthesis in Bacillus sphaericus. Purification and characterization of the pimeloyl-CoA synthase, and uptake of pimelate.